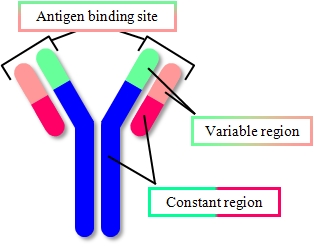
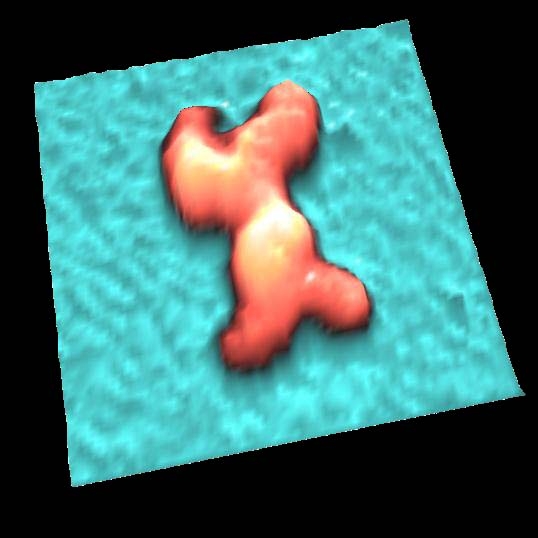
IgG抗体
0.5 frame / sec.(4倍速変換)
IgG 抗体を溶液中で測定しました。
Y字型の構造が鮮明に観察されました。
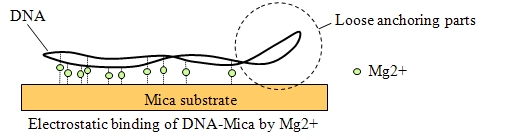
サンプルを基板に強固に固定する必要がなく、
活性のある‘生きた’分子の一瞬の動きを鮮明にとらえることができます。
 |
 |
| IgG抗体のY字構造図 | 高速AFMで測定したIgG抗体 |
プラスミド DNA
従来のAFMではサンプルの固定の弱い箇所は走査中に揺らぐため、画像になりません。
そのために強固な固定が必要になります。
これに対し、高速AFMは揺らぐ前に1画面の走査が終わるため、ゆるく固定したプラスミドDNAも鮮明に測定できます。
強固な固定が不要であるため、サンプルの反応性を損ないません。
EndonucleaseによるDNA分解過程: DNase I
スキャン速度: 1 frame / sec.(30倍速変換)
DNase IはDNAをランダムに切断します。
λDNAが徐々に細片化されていく様子が観察されています。
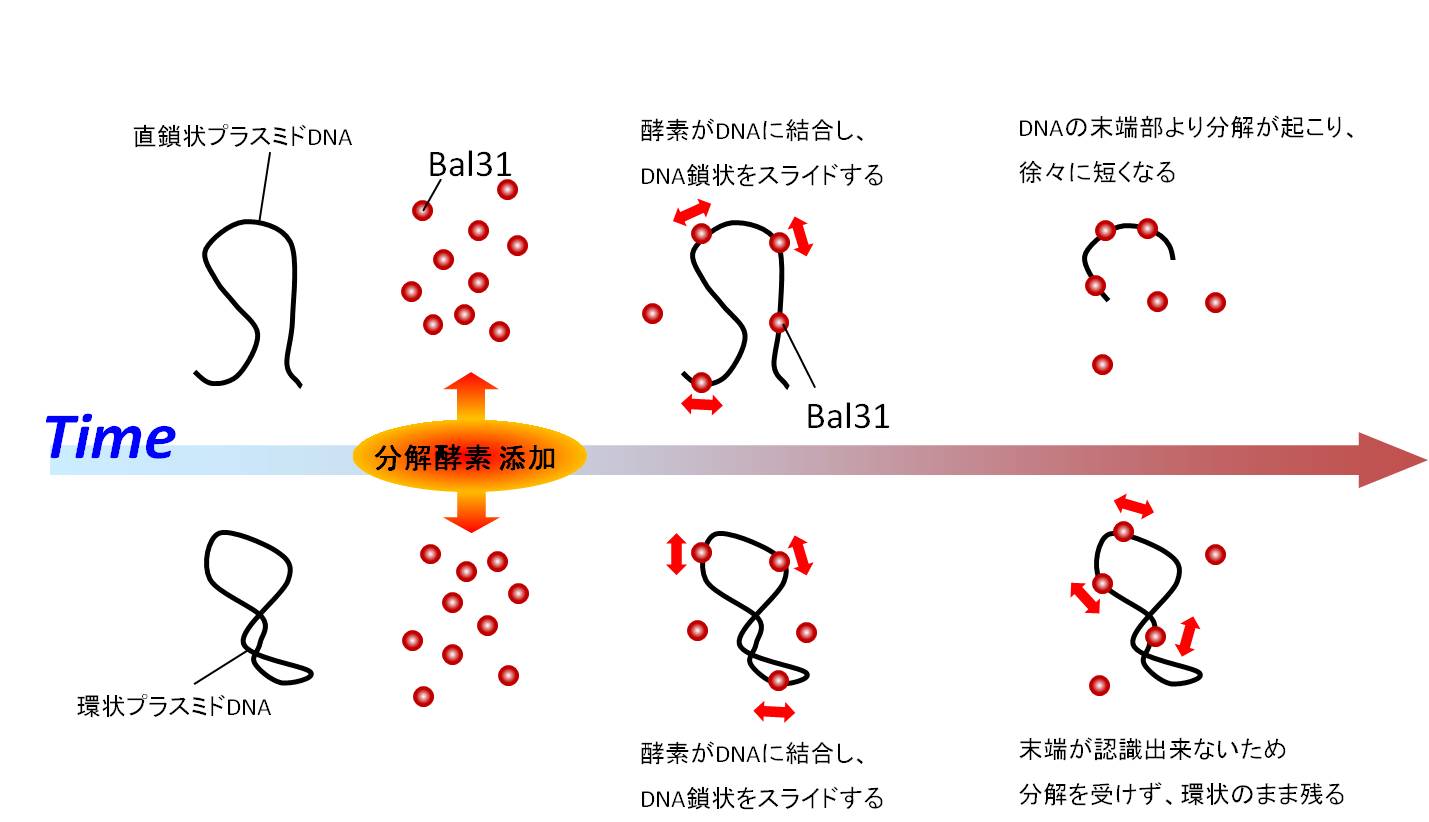
ExonucleaseによるDNA分解過程: Bal31
スキャン速度: 1 frame / sec.(30倍速変換)
粒子状に見えるBal31がPUC18プラスミドDNAの末端から切断していきます。
画面上の直鎖状DNAは徐々に短くなっていきます。画像左下の環状DNAは末端がないため切断を受けません。
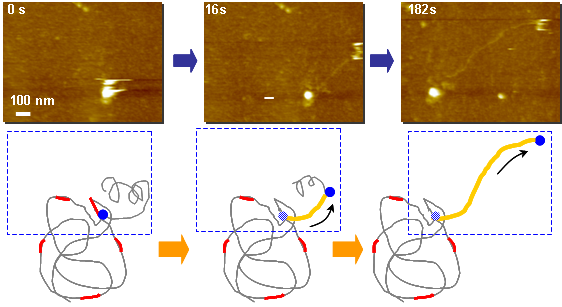
DNA polymerase によるDNA伸長過程: Phi29
Scan Size: 1000 nm × 750 nm
1 frame / sec.(8倍速変換)
鋳型となる1本鎖DNA(λDNA)が基板に固定されています。
Random hexamer primer(赤色図示)が鋳型DNAに結合した箇所を起点にし、
Phi29 Polymerase(青色図示)が基質であるdNTPを利用して相補鎖DNAを合成します。
2本鎖DNA(黄色図示)が時間経過と共に伸長する様子が確認できます。
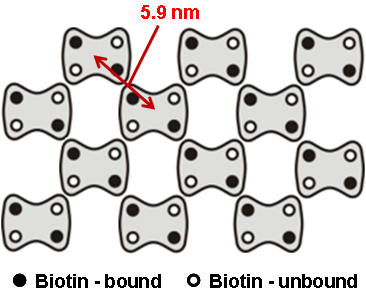
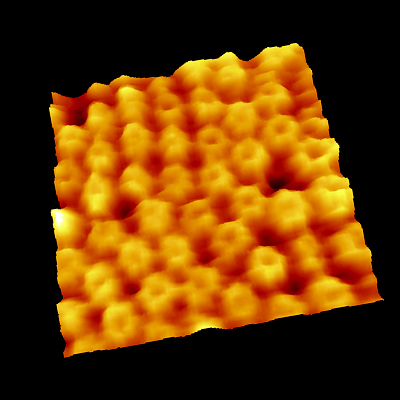
ストレプトアビジン二次元結晶
ビオチンを含む脂質二重膜上にストレプトアビジン二次元結晶を形成し、測定しました。
ストレプトアビジン(大きさ5.9 nm) が規則的に並んでいる様子が観察されています。
脂質膜の流動により点欠陥(穴)が移動していく様子がとらえられています。
[ このページの先頭に戻る ]
 |
 |
|
免疫グロブリンA(IgA抗体)
・Scan Size: 70 nm × 70 nm
|
免疫グロブリンG(IgG抗体)
・Scan Size: 150 nm × 150 nm |
 |
 |
|
ミオシンⅡ
・Scan Size: 500 nm × 500 nm
|
ストレプトアビジン二次元結晶
・Scan Size: 90 nm × 90 nm |
 |
 |
|
GroELシャペロン
・Scan Size: 90 nm × 90 nm
|
バクテリオロドプシン
・Scan Size: 100 nm × 100 nm |
 |
 |
|
脂質膜
・Scan Size: 3500 nm × 3500 nm
|
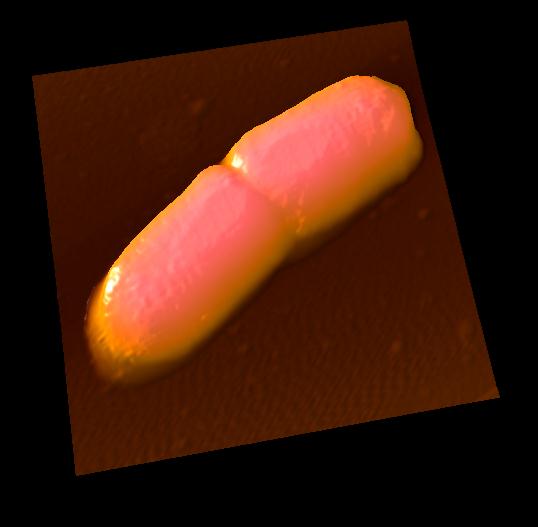
細胞分裂直後の大腸菌
・Scan Size: 3000 nm × 3000 nm |
 |
 |
|
350nm ポリスチレンビーズ
・Scan Size: 3000 nm × 3000 nm
|
350nm ポリスチレンビーズ
・Scan Size: 900 nm × 900 nm |
 |
|
|
プラスミドDNA
・Scan Size: 250 nm × 250 nm
|
|
[ このページの先頭に戻る ]
1. アクチンフィラメントに沿って‘歩く’ミオシンV
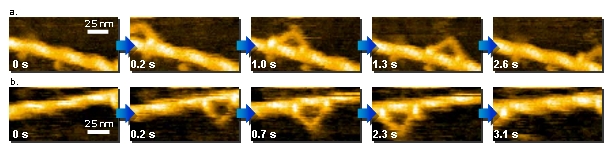
a., b. ミオシンV歩行運動の動画観察 ( a. 130 nm x 65 nm, b. 125 nm x 62.5 nm.)
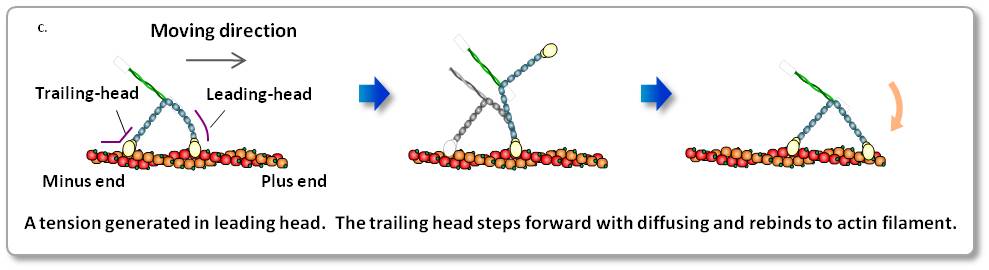
c. ミオシンV歩行運動の模式図
ミオシンVは細胞の骨格をつくるアクチンフィラメント上を移動し、細胞内小器官(オルガネラ)等を運ぶ役割を持っています。
高速AFMによりミオシンVの歩行運動の視覚化を実現し、これまでの研究で明らかにされてきた運動メカニズムを視覚的に証明することに成功しました。
ミオシンVは、その前脚に発生する張力により、回転運動を伴って後脚を前に踏み出します。
さらに、時折足踏みのような運動をすることなど、数々の新しい知見も得ることが出来ました。
N. Kodera et al. Nature 468, 72 (2010). Kanazawa University
2. 光励起によるバクテリオロドプシンの構造変化
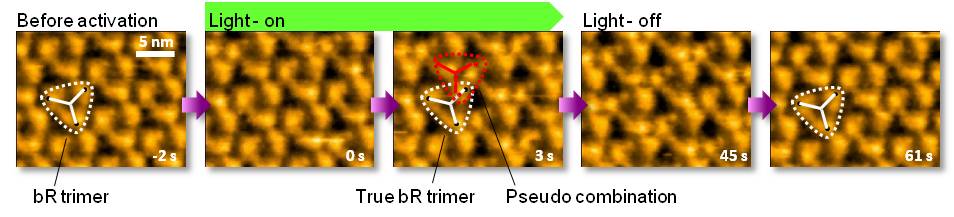
光駆動プロトンポンプタンパク質として知られるバクテリオロドプシン(bR)の光励起による構造変化の動態観察を行いました。
光反応サイクルが野生型bRよりも遅い変異bRを用いて、マイカ基板上に形成された紫膜にグリーンレーザー(532 nm)を照射すると、bR分子は隣り合う三量体のbR分子の方に接近し、あたかも新しい組み合わせの三量体が形成されたかのように観察されます。
この変化は可逆的であり、光照射停止数秒後で元に戻るだけでなく、高い再現性で光応答して繰り返すことが確認されました。
M. Shibata et al. Nature Nanotech. 5, 208 (2010). Kanazawa University
3. ストレプトアビジン二次元結晶
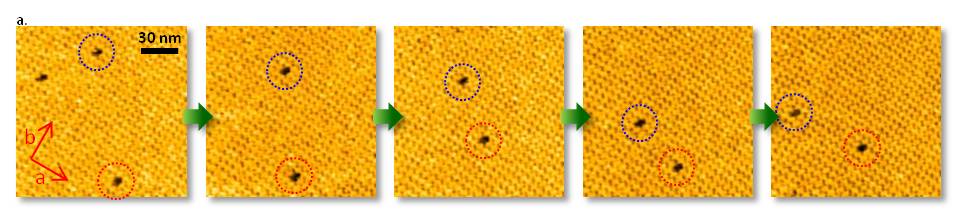
a. ストレプトアビジン二次元結晶の点欠陥の拡散の動画観察
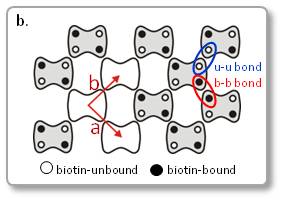
b. ビオチン結合ユニットを示した模式図
ビオチンを含む脂質二重膜上のストレプトアビジン二次元結晶を形成し、結晶格子の点欠陥の拡散を観察しました。
点欠陥の拡散速度は、b軸に沿った拡散の方がa軸に沿った拡散よりも早いことが明らかになりました。
これは、ストレプトアビジン二次元結晶内の各結合サブユニット(u-u結合、b-b結合)の親和性が、u-u結合よりb-b結合の方が高いことを示しています。
このように、結晶内でのタンパク質挙動や、結晶成長メカニズムの議論が可能となります。
D. Yamamoto et al. Nanotechnology 19, 384009 (2008). Kanazawa University
[ このページの先頭に戻る ]
高速原子間力顕微鏡ユーザーによる論文のリストです。
測定の参考などにご利用ください。
[ このページの先頭に戻る ]